
Abstract
This paper addresses the general problem of how to rationally choose an algorithm for phylogenetic inference. Specifically, the controversy between maximum likelihood (ML) and maximum parsimony (MP) perspectives is reframed within the philosophical issue of theory choice. A Kuhnian approach in which rationality is bounded and value-laden is offered and construed through the notion of a Style of Modeling. A Style is divided into four stages: collecting remnant models, constructing models of taxonomical identity, implementing modeling algorithms, and finally inferring and confirming evolutionary trees or cladograms. The identification and investigation of styles is useful for exploring sociological and epistemological issues such as individuating scientific communities and assessing the rationality of algorithm choice. Regarding the last point, this paper suggests that the values motivating ML and MP perspectives are justified but only contextually; these algorithms also have normative force because they can be therapeutic by allowing us to rationally choose among several competing trees, nonetheless this force is limited and cannot be used in order to decide the controversy tout court.
Similar content being viewed by others
Notes
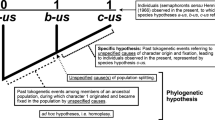
Here I am sidestepping the problem of how to connect the gene-trees (or some other character-laden tree) with the species-trees. Now it is widely accepted that a tree derived from a particular character may not coincide with the species-tree. This might occur, for example, as a result of the Lateral Gene Transfer (LGT). This last phenomenon—LGT—has originated severe skepticism among some practitioners up to the point of actually considering the viability of knowing the real phylogenies of living beings as nothing but a chimerical dream (e.g. Doolittle 1999; Doolittle and Bapteste 2007; but see Woese (2000) for a defense of the viability of inferring phylogenies even granting LGT). In any case, the skepticism seems to be mainly directed against the possibility of interpreting the cladograms as species-trees representations of actual phylogenies; no one seriously doubts the premise that evolution is a singular event, i.e. that evolution occurred only in one way, even if it cannot be represented through a tree and even if its complexity was and is so high that it will never be feasible to actually reconstruct it. My argument requires only to accept that evolution is a singular event and thus, that if it can be represented, then whenever two or more representations on it are discordant at least one of them is not depicting evolution as it actually occurred.
This paper is certainly not the first one that has intended to apply a Kuhnian approach to Systematics, see, for example, Richards (2002) who made a Kuhnian analysis focusing on how parsimony can be interpreted in different ways. He has followed Kuhn (1977a, b, c) article on Objectivity, values and rationality, this article will also be fundamental in this present exposition.
It is worthwhile noting that the problem of the computational limits of the human brain is different from the problem of the amount of information that is available for a subject and the way in which partiality affects the judgment of the subject. Herbert Simon considered that both problems undermined the positivistic conception of an algorithmic (or logically bounded) rationality.
The notion of explanation is one of the thickest philosophical concepts of our time. Here I am using it in its widest form and making no distinction between, for example, ontic, epistemic or modal conceptions about explanation. See Salmon (1990) for a history and a taxonomy of the many ways in which explanation has been understood.
A value, as I am using here, can be conceived as a hypothetical or conditional imperative.
Feminist philosopher Helen Longino (1993, 1995) argues that values such as the attention to human needs should be important for science; this value is especially relevant within the context of modern technologies and problems such as biotechnology and global warming. I believe that in those contexts the importance of a value-laden rationality, in opposition to an instrumental rationality, acquires more relevance. But for the rest of the article I will sidestep this point.
This lemma is inspired by the work of Echeverría (2006) who offers a similar presentation on the thesis of a bounded and value-laden rationality.
Here I use the concept of a space of acceptability in a broad sense. The concept could be further subdivided in the space of stipulability, the space of assertability and the space of acceptability in the narrow sense. The space of stipulability will comprise all of those theories that are composed of sentences (or more generally, of representations) that can have a truth-value (false or true) given the ontological and epistemological assumptions of a given community. The space of assertability will comprise all of those theories that are, in light of a given community with its associated intervening strategies, empirically equivalent. At last, the space of acceptability in the narrow sense will comprise all of those theories that are considered acceptable for particular aims, for example, in order to explain, predict, guide intervention, model a system in some particular conditions, etc. Depending on the aims, the space of acceptability will or will not be a subset of the space of assertability. Both are, nonetheless, subsets of the space of stipulability. These distinctions were inspired by Kuhn (1991) and Wang (2002).
My use of the notion of ‘Style’ is indebted to the philosophical analysis of Winther (2003, 2006, 2009a) who proposed to analyze biology in terms of styles of research; these styles guide the way in which scientists abstract, explain and model. In particular, he identifies two general styles in biology: compositional and formal (Winther 2003, 2006). He developed this idea, in part, by articulating views developed by Ian Hacking, Arnold Davidson and Crombie, among many others.
I will sidestep any possible conflict that may result of these aims. Dupré (2002) forcefully argues that classification should not depend on only one criterion such as monophyly because systematists are not the only users of classifications. I will not delve further into any possible philosophical complication that may arise from this.
Ron Amundson (2005) shows how Systematics—through the notion of ‘homology—is deeply indebted to the Structuralist thought of authors such a Richard Owen. This point is relevant in this context because giving an anatomical (or physiological or behavioral) description is not merely stating what an organism looks like but also to suggest, to hypothesize, to whom it looks alike.
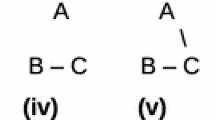
Elliot Sober (2008, Sect. 4.4) shows how categorizing a trait as a binary, multi-state or continuous can affect the amount of information it provides. He also indicates the way in which evolutionary models can probabilify different transformation series and thus modify the final topology of an evolutionary tree. I take these two cases as examples of the importance of this second stage in the modeling process. In the first example, the principle of total evidence demands a categorization that will maximize the amount of information. Nevertheless this principle must produce a categorization with a biological meaning and this is the moment when the partitioning frames start to serve a useful role. In the second example, it could be argued that the very possibility of constructing a transformation series and so of constructing an evolutionary model depends on the partitioning frames employed. This also points to a tendency that, although not a rule, is common in Systematics: likelihoodists utilize transformation series—implicit in their evolutionary models—more often than not and Cladists reject them more often than not.
The partitioning frames are also crucial in the process of analyzing, for example, when a structure can be legitimately subdivided into smaller parts that later will be codified as many characters with their associated character states. If we lack of any grounds for making such a subdivision will be in practice weighting a character and so biasing the outcome of the phylogenetical analysis.
A counter-value is a disbelief on the usefulness of some particular criterion that in principle is offered as a manner for discriminating particular ways of achieving an aim.
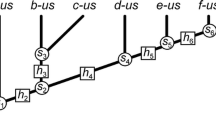
To differentiate the models produced in the four stages by MP and ML I will circumscribe the term ‘cladogram’ to the trees constructed by MP and the term ‘evolutionary tree’ as the trees constructed by ML. Cladograms only have topologies while Evolutionary trees have topologies and geometries.
But, to the best of my knowledge, the models that incorporate molecular and morphological traits are usually employed by Bayesians and not by Likelihoodists.
What does it mean to explain something in Systematics is a debated issue, see, for example, Bock (2004).
By Therapeutic capacity I understand the ability of a norm to provide us with some prescriptive, mandatory regulations, which stipulate the kinds of acceptable outcomes or actions that can be performed according to this norm. For example, how a given aim can be pursued or whether the inferences leading to a conclusion are the result of a reliable mechanism. It should be clear that these prescriptive regulations are hypothetical imperatives.
A historical précis might be relevant here. Olivier Rieppel (2003, 2005) criticized the use of a popperian framework in phylogenetics. I do not address his criticisms because I believe that rational disagreements are historically and contextually circumscribed. Thus, after the criticisms presented by Rieppel, the systematists run the risk of passing from a rational disagreement into an irrational disagreement if they simple ignore Rieppel’s criticisms. However, given the fact that my aim is not to analyze the validity of the Popperian framework in Systematics but only to scrutinize how some criterion is interpreted in different ways by different communities, it is not directly relevant, but not for that reason uninteresting by itself, to discuss the validity of that criterion.
Among those other factors is the incapacity of ML to include non-molecular characters. Many Cladists see this as simply ignoring many informative characters, and in the process a part of the phylogenetic signal is lost. In the following I will set aside this issue but I want to emphasize that in a more complete analysis it should certainly be included. Another related problem will be the NP-completeness or the impossibility of finding within an algorithmic time a global optimum through an exhaustive exploration of the tree-space; for many Cladists ML will be twice NP-complete because it must calculate distances and not just seek for topologies as MP does. This means that ML has a much bigger tree-space than MP and so the chance of finding the right tree is diminished.
Lyons-Weiler et al. (1997) proposed a strategy that did not assume any evolutionary model whatsoever and so, according to these authors, could help us to detect when we are in the Felsenstein zone. If this strategy (or any other technique that does not assume a particular evolutionary model) could be proved successful, then the rational disagreement diagnosed here would have to be reevaluated.
Richards (2002) has made a similar point and he also utilizes a Kuhnian approach.
References
Amundson R (2005) The changing role of the embryo in evolutionary thought: structure and synthesis. Cambridge University Press, Cambridge
Anderson FE, Swofford DL (2004) Should we be worried about long-branch attraction in real data sets? Investigations using metazoan 18S rDNA. Mol Phylogenet Evol 33:440–451. doi:10.1016/j.ympev.2004.06.015
Bergsten J (2005) A review of long-branch attraction. Cladistics 21:163–193. doi:10.1111/j.1096-0031.2005.00059.x
Bock WJ (2004) Explanation in systematics. In: Williams DM, Forey PL (eds) Milestones in systematics. CRC Press, Boca Raton
Brower AVZ (2000) Evolution is not a necessary assumption of cladistics. Cladistics 16:143–154. doi:10.1111/j.1096-0031.2000.tb00351.x
Buckley T (2000) New perspectives on parsimony and likelihood. Trends Ecol Evol 15(9):356. doi:10.1016/S0169-5347(00)01945-5
De Queiroz K, Poe S (2001) Philosophy and phylogenetic inference: a comparison of likelihood and parsimony methods in the context of Karl Popper’s writings on corroboration. Syst Biol 50(3):305–321. doi:10.1080/106351501300317941
Doolittle WF (1999) Phylogenetic classification and the universal tree. Science 284(5423):2124–2128. doi:10.1126/science.284.5423.2124
Doolittle WF, Bapteste E (2007) Pattern pluralism and the tree of life hypothesis. Proc Natl Acad Sci USA 104(7):2043–2049. doi:10.1073/pnas.0610699104
Dupré J (2002) Humans and other animals. Oxford University Press, Oxford
Echeverría J (2006) Dos Dogmas del racionalismo (y una propuesta alternativa) (Manuscript)
Faith DP (2006) Science and philosophy for molecular systematics: which is the cart and which is the horse? Mol Phylogenet Evol 38(2):553–557. doi:10.1016/j.ympev.2005.08.018
Faith DP, Trueman JWH (2001) Towards an inclusive philosophy for phylogenetic inference. Syst Biol 50(3):331–350. doi:10.1080/106351501300317969
Farris JS, Källersjö M, Albert VA, Allard M, Anderberg A, Bowditch B, Bult C, Carpenter JM, Crowe TM, De Laet J, Fitzhugh K, Frost D, Goloboff P, Humphries CJ, Jondelius U, Judd D, Karis PO, Lipscomb D, Luckow M, Mindell D, Muona J, Nixon K, Presch W, Seberg O, Siddall ME, Struwe L, Tehler A, Wenzel J, Wheeler Q, Wheeler W (1995) Explanation. Cladistics 11:211–218. doi:10.1111/j.1096-0031.1995.tb00086.x
Felsenstein J (1978) Cases in which parsimony or compatibility methods will be positively misleading. Syst Zool 27:401–410. doi:10.2307/2412923
Felsenstein J (1981) Evolutionary trees from DNA sequences: maximum likelihood approach. J Mol Evol 17:368–376. doi:10.1007/BF01734359
Feyerabend PK (1970) Consuelos para el especialista. In: Lakatos I, Musgrave A (eds) La crítica y el desarrollo del conocimiento. Ediciones Grijalbo, España, 1975
Fitzhugh K (2006) The philosophical basis of character coding for the inference of phylogenetic hypotheses. Zool Scr 35(3):261–286. doi:10.1111/j.1463-6409.2006.00229.x
Griesemer J (1990) Modeling in the museum: on the roleof remnant models in the work of Joseph Grinell. Biol Philos 5:3–36. doi:10.1007/BF02423831
Hacking I (1983) Representing and intervening: introductory topics in the philosophy of natural science. Cambridge University Press, Cambridge
Helfenbein KG, DeSalle R (2005) Falsifications and corroborations: Karl Popper’s influence on systematics. Mol Phylogenet Evol 35:271–280. doi:10.1016/j.ympev.2005.01.003
Hennig W (1966) Phylogenetic systematics. University of illinois Press, Urbana
Huelsenbeck JP (1997) Is the Felsenstein zone a fly trap? Syst Biol 46:69–74. doi:10.2307/2413636
Hull D (1988) Science as a process. University of Chicago Press, Chicago
Kluge AG (1999) The science of phylogenetic systematics: explanation, prediction, and test. Cladistics 15:429–436. doi:10.1111/j.1096-0031.1999.tb00279.x
Kluge AG (2001) Parsimony with and without scientific justification. Cladistics 17:199–210. doi:10.1111/j.1096-0031.2001.tb00117.x
Kuhn TS (1969) La estructura de las Revoluciones científicas (Postdata). FCE, México
Kuhn TS (1971) Las revoluciones como cambios de la concepción del mundo. In: Pérez Ransanz AR, Olivé L (eds) Filosofía de la Ciencia: Teoría y observación. Siglo XXI-UNAM, México
Kuhn TS (1977a) Objectivity, value judgment, and theory choice in the essential tension. The University of Chicago Press, Chicago
Kuhn TS (1977b) Second thoughts on paradigms. In: Kuhn TS (ed) The essential tension. The University of Chicago Press, Chicago
Kuhn TS (1977c) Metaphor in science. In: Conant J, Haugeland J (eds) The road since structure. The University of Chicago Press, Chicago
Kuhn TS (1983a) Conmensurabilidad, comparabilidad y compatibilidad in ¿Qué son las Revoluciones Científicas? y otros ensayos. Paidós, España, 1989
Kuhn TS (1983b) Racionalidad y elección de Teorías in ¿Qué son las Revoluciones Científicas? y otros ensayos. Paidós, España, 1989
Kuhn TS (1987) ¿Qué son las revoluciones científicas? in ¿Qué son las Revoluciones Científicas? y otros ensayos. Paidós, España, 1989
Kuhn TS (1991) The road since structure. In: Conant J, Haugeland J (eds) The road since structure. The University of Chicago Press, Chicago
Longino HE (1990) Science as social knowledge: values and objectivity in scientific inquiry. Princeton University Press, Princeton
Longino HE (1993) Subjects, power and knowledge: descriptions and prescriptions in feminist philosophies of science. In: Alcoff L, Potter E (eds) Feminist epistemologies. Routledge, London
Longino HE (1995) Gender. Political Theor Virtues Synth 104:383–397
Longino HE (2000) Toward an epistemology for biological pluralism. In: Creath R, Maienschein J (eds) Biology and epistemology. Cambridge University Press, Cambridge, pp 261–286
Longino HE (2002) The fate of knowledge. Princeton University Press, Princeton
Lyons-Weiler J, Guy A, Hoelzer GA (1997) Escaping from the Felsenstein zone by detecting long branches in phylogenetic data. Mol Phylogenet Evol 8(3):375–384. doi:10.1006/mpev.1997.0450
Pérez Ransanz AR (1999) Kuhn y el cambio científico. FCE, México
Popper K (1962) La Lógica de la Investigación Científica. In: Tecnos (ed) Madrid
Popper K (1963) Conjectures and refutations: the growth of scientific knowledge. Routledge, London
Rescher N (1993) La Racionalidad. Tecnos, Spain
Richards R (2002) Kuhnian values and cladistic parsimony. Perspect Sci 10:1–27. doi:10.1162/106361402762674780
Rieppel O (2003) Popper and systematics. Syst Biol 52(2):259–271. doi:10.1080/10635150390192762
Rieppel O (2005) A skeptical look at justification. Cladistics 21:203–207. doi:10.1111/j.1096-0031.2005.00057.x (Letter to the Editor)
Rokas A (2001) Getting it right for the wrong reason. Trends Ecol Evol 16(12):668. doi:10.1016/S0169-5347(01)02376-X
Salmon W (1990) Four decades of scientific explanation. Pittsburgh University Press, Pittsburgh
Siddall ME (1998) Success of parsimony in the four-taxon case: long-branch repulsion by likelihood in the Farris zone. Cladistics 14:209–220. doi:10.1111/j.1096-0031.1998.tb00334.x
Siddall ME (2001) Philosphy and phylogenetic inference: a comparison of likelihood and parsimony methods in the context of Karl Popper’s writings on corroboration. Cladistics 15:17 (Letter to the Editor)
Siddall ME, Kluge AG (1997) Probabilism and phylogenetic inference. Cladistics 13:313–336. doi:10.1111/j.1096-0031.1997.tb00322.x
Siddall ME, Whiting MF (1999) Long-branch abstractions. Cladistics 15:9–24. doi:10.1111/j.1096-0031.1999.tb00391.x
Simon H (1997) Models of bounded rationality. The MIT Press, Cambridge
Sober E (1988) Reconstructing the past: parsimony, evolution and inference. The MIT Press, Cambridge
Sober E (2004) The contest between parsimony and likelihood. Syst Biol 53(4):644–653. doi:10.1080/10635150490468657
Sober E (2008) Evidence and evolution. The logic behind the science. Cambridge University Press, Cambridge
Wang X (2002) Taxonomy, truth-value gaps and incommensurability: a reconstruction of Kuhn’s taxonomic interpretation of incommensurability. Stud Hist Philos Sci 33:465–485. doi:10.1016/S0039-3681(01)00039-5
Wimsatt WC (1972) Teleology and the logical structure of function statements. Stud Hist Philos Sci 3:1–80. doi:10.1016/0039-3681(72)90014-3
Winther RG (2001) Varieties of modules: kinds, levels, origins, and behaviors. J Exp Zool 291:116–129. doi:10.1002/jez.1064
Winther RG (2003) Formal biology and compositional biology as two kinds of biological theorizing. Dissertation, History and Philosophy of Science Department, Indiana University
Winther RG (2006) Parts and theories in compositional biology. Biol Philos 21:471–499. doi:10.1007/s10539-005-9002-x
Winther RG (2009a) Teorías, Prácticas y Estilos de Investigación Científica. In: Martínez S, Huang X, Guillaumin G (eds) Filosofía de las prácticas científicas. Hacia una filosofía de la ciencia no centrada en teorías. Universidad Autónoma Metropolitana, México (accepted)
Winther RG (2009b) Character analysis in cladistics: abstraction, reification, and the search for objectivity. Acta Biotheor. doi:10.1007/s10441-008-9064-7
Woese CR (2000) Interpreting the universal phylogenetic tree. Proc Natl Acad Sci USA 97(15):8392–8396. doi:10.1073/pnas.97.15.8392
Acknowledgments
First and foremost I want to thank Rasmus Winther and Paco Vergara. They have been, are, and surely will remain as great influences in my life. They have been my teachers and also my friends. Thanks for inviting me to join this particular project as well. I also want to thank Martha Martínez and Helga Ochoterena, who have also taught me a lot about Systematics and motivated, by doing that, many of the points here presented; any failure is of course only mine. I would also like to thank Gabriela Guerrero’s technical assistance when writing this paper. And lastly, I want to thank those philosophers who have greatly influenced my thinking: Ana Rosa Pérez Ransanz (How could I possibly understand Kuhn without your guidance), Sergio Martínez and Ángeles Eraña (your reflections on rationality have been an inspiration), and finally I want to thank Carlos López Beltrán for teaching me how history matters even in the realm of reason.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mc Manus, F.G. Rational Disagreements in Phylogenetics. Acta Biotheor 57, 99–127 (2009). https://doi.org/10.1007/s10441-009-9072-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10441-009-9072-2