
Abstract
My aim here is to show that approximate truth as a paraconsistent notion (neutral to the realism/anti-realism debate) can be successfully incorporated into the analysis of scientific unification, thus advancing towards a more realistic representation of theory development that takes into account the controversies that often loom alongside the progress of research programmes. I support my analysis with a case study of the recent debate in ecology centred around the existence of the paradox of enrichment and the controversy between ecological models of predation that employ prey-dependent and ratio-dependent functional responses. These models were initially proposed as equally good representations of the basic aspects of predator–prey dynamics. However, both models generated inconsistent observational consequences and, therefore, introduced a contradiction within predator–prey theory. I argue that by accepting these models as approximately true representations of predator–prey dynamics we can convey how the available observational data have been successfully systematized in a consistent way under them. This first step in resolving the controversy relied on building a series of contrastive arguments based on both models’ derivations about population dynamics and the available empirical data. The heightening of this contrast between the models, in turn, was also essential in defining a limiting function which can be used to integrate both models and reach a new unified expression of predator–prey dynamics.
Similar content being viewed by others


Notes
I.e., predator–prey interaction.
The functional response in predator–prey modeling is the function which governs the intake rate of the predator in relation with the food density.
A basic assumption that I adopt for this paper is that we are dealing with ordinary cases and not dialetheias (Priest 2006).
See the “Appendix” for some of the open issues in defining the functional response and the resulting models of predator–prey dynamics.
Philosophers of science and logicians are advised to note that the paradox of enrichment does not correspond to the usual meaning of a paradox as a contradictory statement. Here, it simply designates a counterintuitive outcome. Normally, we would expect that if the conditions of the environment permit an increase in the population of prey, the whole system will respond with proportional growth of both populations, and no extinction event is likely to occur.
In some lakes, for instance, the increase in phytoplankton variability has been related to nutrient enrichment (Cottingham et al. 2000).
Direct interference is the situation in which the functional response is affected at high predator densities because predators compete for the available prey. Indirect interference affects the functional response again at high predator densities, not because of direct interaction between predators but because of prey depletion.
Taking into consideration the density of the predator population for determining the functional response became known as predator-dependent functional response (Abrams and Ginzburg 2000).
A further point that justifies my choice not to apply C&P is that, whenever C&P structures have an empty permeability function (or no information flow between the chunks), they reduce to Schotch and Jennings's non-adjunction strategy (Brown and Priest 2004), which is at the heart of Brown's approach.
For a more complete exposition of the features of Brown's paraconsistent consequence relation see Consequence as Preservation: Some Refinements (Brown 2013).
A further point is that, by applying the prey-dependent functional response, the paradox of enrichment becomes a situation that must be accounted for. Under the model’s conceptualization of functional response, the counterfactual problem of “Why is the paradox of enrichment not observed?” becomes a legitimate explanatory-seeking question. It is expected that studies proposing at explanandum factors will pile up. Given a formal contrast with the ratio-dependence model, the paradox of enrichment can be more easily assessed as a theoretical artifact.
The consequence relations within chunks are expected to be the familiar consistency-preserving/satisfiability preserving relations of standard-issue logics (both classical and weaker, such as intuitionist).
References
Abrams, P. A. (1994). The fallacies of ratio-dependent predation. Ecology, 75, 1842–1850.
Abrams, P. A. (2014). Why ratio dependence is (still) a bad model of predation. Biological Reviews, 90, 794–814.
Abrams, P. A. (2015). Why ratio dependence is (still) a bad model of predation. Biological Reviews Cambridge Philosophy of Society, 90, 794–814.
Abrams, P. A., & Ginzburg, L. (2000). The nature of predation: Prey-dependent, ratio-dependent or neither? Trends in Ecology and Evolution, 15, 337–341.
Arditi, R., & Akcakaya, H. R. (1990). Underestimation of mutual interference of predators. Oecologia, 83, 358–361.
Arditi, R., & Ginzburg, L. (1989). Coupling in predator–prey dynamics: Ratio-dependence. Journal of Theoretical Biology, 139(3), 311–326.
Arditi, R., & Ginzburg, L. (2012). How species interact: Altering the standard view on trophic ecology. New York: Oxford University Press.
Bangu, S. (2016). Scientific explanation and understanding: Unificationism reconsidered. European Journal for Philosophy of Science. https://doi.org/10.1007/s13194-016-0148-y.
Barraquand, F. (2014). Functional responses and predator–prey models: A critique of ratio dependence. Theoretical Ecology, 7, 3–20.
Bartelborth, T. (2002). Explanatory unification. Synthese, 130, 91–107.
Beddington, J. (1975). Mutual interference between parasites or predators and its effect on searching efficiency. Journal of Animal Ecology, 44, 331–340.
Benham, R., Mortensen, C., & Priest, G. (2014). Chunk and permeate III: The Dirac delta function. Synthese, 191, 3057–3062.
Brauer, F., & Castillo-Chavez, C. (2012). Mathematical models in population biology and epidemiology (Vol. 2). New York: Springer.
Brown, B. (2002). Approximate truth: A paraconsistent account. In J. Meheus (Ed.), Inconsistency in science (pp. 81–103). Dordrecht: Kluwer Academic Publishers.
Brown, B. (2013). Consequence as preservation: Some refinements. In F. Berto, E. Mares, K. Tanaka, & F. Paoli (Eds.), Paraconsistency: Logic and applications (pp. 123–139). Springer.
Brown, B. (2015). Peter Vickers: Understanding inconsistent science (book review). Journal for General Philosophy of Science, 46(2), 413–418.
Brown, B., & Priest, G. (2004). Chunk and permeate: A paraconsistent inference strategy-part 1—The infinitesimal calculus. The Journal of Philosophical Logic, 33, 379–388.
Brown, B., & Priest, G. (2008). Chunk and permeate II: Weak aggregation, permeation and old quantum theory. Melbourne: Fourth World Congress on Paraconsistency.
Chowell, G., & Viboud, C. (2016). Is it growing exponentially fast? Impact of assuming exponential growth for characterizing and forecasting epidemics with initial near-exponential growth dynamics. Infectious Disease Modelling, 1(1), 71–78.
Cottingham, K. L., Rusak, J. A., & Leavitt, P. R. (2000). Increased ecosystem variability and reduced predictability following fertilisation: Evidence from palaeolimnology. Ecology Letters, 3, 340–348.
Davey, K. (2014). Can good science be logically inconsistent? Synthese, 191, 3009–3026.
DeAngelis, D. L., Goldstein, R. A., & O’Neill, R. V. (1975). A model for trophic interaction. Ecology, 56, 881–892.
Friedman, M. (1974). Explanation and scientific understanding. Journal of Philosophy, 71, 5–19.
Ginzburg, L., & Colyvan, M. (2004). Ecological orbits: How planets move and populations grow. New York: Oxford University Press.
Ginzburg, L., & Jensen, C. (2008). From controversy to consensus: The indirect interference functional response. Verhandlungen der InternationalenVereinigung für Theoretische und Angewandte Limnologie, 30, 297–301.
Hassell, M. P., & Varley, G. C. (1969). New inductive population model for insect parasites and its bearing on biological control. Nature, 223, 1133–1137.
Holling, C. S. (1959). The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. The Canadian Entomologist, 91(5), 293–320. https://doi.org/10.4039/Ent91293-5.
Holt, R. D. (2011). Natural enemy-victim interactions: Do we have a unified theory yet? In S. M. Scheiner & M. R. Willig (Eds.), The theory of ecology (pp. 125–161). Chicago: University of Chicago Press.
Jensen, C. (2008). Predation and its consequences: Insights into the modeling of interference. Stony Brook Theses & Dissertations [SBU], Stony Brook University. https://dspace.sunyconnect.suny.edu/handle/1951/44258. Accessed 09 May 2017.
Jost, C. (1998). Comparing predator–prey models qualitatively and quantitatively with ecological time-series data. Paris: Institut national agronomique Paris-Grignon.
Kitcher, P. (1989). Explanatory unification and the causal structure of the world. In P. Kitcher & W. Salmon (Eds.), Scientific explanation (pp. 410–505). Minneapolis: University of Minnesota Press.
Laudan, L. (1981). A confutation of convergent realism. Philosophy of Science, 48, 19–48.
Morrison, M. (2000). Unifying scientific theories. Cambridge: Cambridge University Press.
Petkov, S. (2015). Explanatory unification and conceptualization. Synthese, 192, 3695–3717.
Priest, G. (2002). Inconsistency and the empirical sciences. In J. Meheus (Ed.), Inconsistency in science (pp. 119–128). Dordrecht: Kluwer Academic Publishers.
Priest, G. (2006). In contradiction. Oxford: Clarendon Press.
Prokopenko, M. C., Turgeon, K., & Fryxell, J. M. (2017). Evaluation of alternative prey-, predator-, and ratio-dependent functional response models in a zooplankton microcosm. Canadian Journal of Zoology, 95, 177–182.
Psillos, S. (2017). World-involving scientific understanding. Balkan Journal of Philosophy, 9(1), 5–18.
Psillos, S. (2018). Realism and theory change in science. In Zalta, E. N. (Ed.), The Stanford Encyclopedia of Philosophy, Summer 2018 Edition. https://plato.stanford.edu/archives/sum2018/entries/realism-theory-change/. Accessed 26 August 2018.
Rosenzweig, M. L. (1971). Paradox of enrichment: Destabilization of exploitation ecosystems in ecological time. Science, 171, 385–387.
Schotch, P. K., & Jennings, R. E. (1980). Inference and necessity. Journal of Philosophical Logic, 9, 327–340.
Schurz, G. (1999). Explanation as unification. Synthese, 120, 94–114.
Shovonlal, R., & Chattopadhyay, J. (2007). The stability of ecosystems: A brief overview of the paradox of enrichment. Journal of Biosciences, 32(2), 421–428. https://doi.org/10.1007/s12038-007-0040-1.
Slavov, N., et al. (2014). Constant growth rate can be supported by decreasing energy flux and increasing aerobic glycolysis. Cell Reports, 7(3), 705–714.
Tyson, R., Haines, S., & Hodges, K. (2010). Modelling the Canada lynx and snowshoe hare population cycle: The role of specialist predators. Theoretical Ecology, 3(2), 97–111.
Tyutyunov, Y., Titova, L., & Arditi, R. (2008). Predator interference emerging from trophotaxis in predator–prey systems: An individual-based approach. Ecological Complexity, 5, 48–58.
Vickers, P. (2013). Understanding inconsistent science. Oxford: Oxford University Press.
Vickers, P. (2014). Theory flexibility and inconsistency in science. Synthese, 191, 2891–2906.
Volterra, V. (1926a). Fluctuations in the abundance of a species considered mathematically. Nature, 118, 558–560.
Volterra, V. (1926b). Variazioni e fluttuazioni del numerod’individui in specie animaliconviventi, Memorie della R. Accademia Nazional edei Lincei, 2, 5–112.
Weber, E., & Van Dyck, M. (2002). Unification and explanation: A comment on Halonen and Hintikka, and Schurz. Synthese, 131, 145–154.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
1.1 Is the Chunk and Permeate Model Applicable to the Functional Response Debate?
My analysis of model controversies and approximate truth relied upon the application of the non-adjunctive approach to paraconsistency. This implicitly required that the observational domain of the initially internally inconsistent theory can be clearly divided between the conflicting models. Two related questions then remain:
-
1.
Is such a rigid contextualization possible?
-
2.
Are there cases where inconsistent models are used jointly to derive information about the same target?
Addressing these problems must be done with some caution. At least for the case of analysing predator–prey theory, this matter cannot be settled with complete certainty. The ecological modelling of food chains and trophic interaction is incomplete and has an ongoing research program (Abrams 2015). The existing literature so far presents good evidence that a clear contrast between ratio and prey dependent functional responses can be made and this contrast has rendered these models virtually exclusive (Abrams and Ginzburg 2000). However, as I noted, prey-dependent and ratio-dependent functional responses stand at the two opposite ends of a spectrum of possible functional responses where various predator dependent models lie as a middle ground (see Table 1). The hypothesis of gradual interference, in turn, can serve as a notion organizing the hierarchy of predator prey models (Holt 2011).
However it is conceivable that a representational overlap might appear between prey-dependent models and some predator-dependent models or between ratio-dependent models and predator-dependent models. I was, however, unable to pinpoint a case where two such models are used jointly, without a clear cut delineation of their applicability. Even in cases where multiple models are employed by independent studies investigating the same target and these models were generating equally good representations of the empirical data, an argument based on their idealized contrastive features has been made which strived to justify the usage of only one model of functional response per particular empirical case.
In order to address the issue of such a representational overlap I will first present the chunk and permeate structure (C&P)—an extension of the non-adjunctive analysis of paraconsistency, which aims at handling such cases. I will also go over three illustrative examples from ecology that can serve as evidence that ecologists are aiming to avoid such overlaps.
Based on this, two conclusions can be drawn. First, the non-adjunctive approach is in many cases sufficient to analyse the reasoning with conflicting idealized models, and secondly that even if a representational overlap is present, the non-adjunctive analysis can be easily extended to accommodate these situations by extending it to a C&P structure.
The C&K structures operate over the non-adjunctive systems of Schotch and Jennings (1980) by adding a permeation filter. This filter permits controlled information flow between the inconsistent fragments. The C&P can be interpreted as a framework for analysis of cases where consistent information about a given target can be derived from jointly inconsistent models. As such, the C&P is inferentially more powerful than the non-adjunctive approach I have used. Formally, C&P can be briefly sketched as follows (Brown and Priest 2004, 2008; Benham et al. 2014):
The first step in defining a C&P structure is equivalent to the fragmenting mechanism of particularizing an inconsistent set of sentences introduced in Sect. 4. For ∑ a set of sentences in a language L, an n-covering of ∑ is an indexed set of sets of sentences σi, 0 ≤ i ≤ n such that \(\cup_{\upsigma i, 0 \le i \le n } \left( {\upsigma i} \right) = \sum .\) If ∑ contains a contradiction, then the chunking strategy to consistent subsets should be used to particularize Σ so that each cell σi is internally consistent.
The second step in defining the C&P structure is introducing a ρ permeability relation which will permit some information transfer between the resulting cells. A C&P structure on Σ,

, is a 3-tuple \(\left\langle {P,\rho ,i_{0} } \right\rangle\) where:
-
1.
P is a consistent covering of ∑, with elements σ1,… σn
-
2.
ρ is a permeability relation, a function from pairs of integers I × I to subsets of L.
-
3.
i0 is the label of the designated chunk, where the conclusions are drawn.
Over these two steps we can then introduce a consequence relation relative to C&P, which will be a result of a series of closure and permeation steps. If σi is the ith cell of P, \(\upsigma_{i}^{n}\) is defined by recursion:
The C&P works by first closing the cells under a consistency preserving consequence relation.Footnote 13 Secondly any sentences in \(\rho \left( {j,i} \right)\) that appear in cell j are added to cell i. Over this we can define a general C&P consequence of Σ:

Note that the C&P structure does not specify which covering of ∑ is the right one, nor does it specify the nature of the permeation relation. Both the covering and the permeation relation thus should be defined in a pragmatic way so as to guarantee the specific inferences that we want to preserve. If the C&P is conducted to the case of jointly inconsistent models which can be used to derive information about the same target the resulting requirements for these models boil down to:
-
1.
The models should be internally consistent—no singular model should lead to a contradictory conclusion about its target;
-
2.
Given some pragmatically defined ρ relation, the separate conclusions drawn from these models can be used jointly so long as the resulting set of conclusions about the target is not itself inconsistent.
The C&P then can cover the rezoning with theories whose formal models are jointly inconsistent but have a partial representational overlap. However, the functional response debate so far does not offer such an example. Three examples will serve as evidence of this fact.
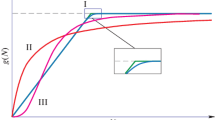
The first one is related to a case where the target displays complexity which might require two models to be used jointly in order to represent it accurately. Such a case might be found in analysis of food chains. However, both ratio-dependent and prey-dependent models have very different predictions for the response of food chains, for instance given an enrichment scenario at the bottom level. According to the standard prey-dependent theory, in reference to the increase in primary production, the responses of the populations strongly depend on their level and on the total number of trophic levels, whilst in ratio-dependent modeling the response will be proportional in all levels. For systems with 2–5 levels with an apex predator, the response to enrichment for prey dependent and ratio dependent models has the structure given in Fig. 1.

Prey Dependence and Ratio Dependence responses in food chains, with enrichment in primary production N, with an apex predator at the top P4 (Arditi and Ginzburg 2012, 63)
Figure 1 provides a clear qualitative contrast between these models; thus whenever they are used for describing food chains, such quantitative differences are sought in order to compare the representational fidelity of these models with the empirical data. Obviously, the empirical data for food chains is incomplete and its complexity leads to fuzziness but the researchers nevertheless are aiming at justifying the usage of one particular model (Arditi and Ginzburg 2012; Abrams 2015).
The second option is to instead seek a case where the models with different functional response are used to represent a particular system but none of them outperforms the other, which then might require that the information about the system should be jointly provided by both of the models. What is most common in the literature, however, is that whenever these models are used to represent the same system, a comparative contrast of the type exemplified in Sect. 4 is used. A recent study compared several functional responses including prey- and ratio-dependent ones (see Table 1) in relation to the interference hypothesis using an experimental setting with the rotifer Brachionus calyciflorus as a prey and the ostracods Heterocypris incongruens as predators (Prokopenko et al. 2017). The researchers found good evidence for the interference hypothesis, which leads models employing predator-dependent functional response and ratio-dependent functional response to outperform prey-dependent models. However, the ratio-dependent model outperformed the other predator-dependent models. Evidence for predator interference as a factor relevant to the functional response was also detected.
Finally, for the cases where the same population dynamics are represented equally well by ratio-dependent and other predator dependent models such as the population cycle of lynxs and snowshoe hares in Canada (Tyson et al. 2010), the ecologists nevertheless strive to find points where one of the models outperforms the other (Barraquand 2014).
These cases I believe present at least partial evidence that the chunking strategy (without permeability) is at play in organizing the hierarchy of predator–prey models within trophic interaction theory.
Rights and permissions
About this article

Cite this article
Petkov, S. Studying Controversies: Unification, Contradiction, Integration. J Gen Philos Sci 50, 103–128 (2019). https://doi.org/10.1007/s10838-018-9431-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10838-018-9431-2