
Abstract
In this paper, I argue (contra some recent philosophical work) that an objective distinction between natural selection and drift can be drawn. I draw this distinction by conceiving of drift, in the most fundamental sense, as an individual-level phenomenon. This goes against some other attempts to distinguish selection from drift, which have argued either that drift is a population-level process or that it is a population-level product. Instead of identifying drift with population-level features, the account introduced here can explain these population-level features based on a property that I label driftability. Additionally, this account shows that biology’s “first law”—the Principle of Drift (Brandon, J Phil 102(7):319–335 2006)—is not a foundational law, but is a consequence of driftability.
Similar content being viewed by others

Notes
These do not exhaust the possibilities. For example, Bouchard and Rosenberg (2004) place drift in the initial conditions, “drift—the departure of these initial conditions from equality in proportion among all physically possible initial conditions” (p. 352); Pfeifer (2005) understands drift to be both a cause of evolution and a result of ignoring or abstracting environmental features; Abrams (2007) takes (one kind of) drift to be understood in terms of the properties of population-level probability distributions: “Natural selection is the aspect of the distribution which is under the control of fitness differences, and drift is the aspect of the distribution which is under the control of population size” (p. 677). I do not have the space in this essay to challenge all of these and other positions. It should be clear in what follows, however, that the position argued for here is not identical to any of these positions.
This view, it should be noted, is not one Rosenberg currently holds.
They note that while this is true of historical reconstructions, such is not the case for “evolutionary theory, which abstracts away from individual causes” (2002, p. 64).
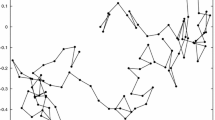
See Fig. 1 from Brandon (2005, p. 162).
See also Brandon and Carson (1996).
Though the positions of Brandon and Walsh et al. differ quite radically in other ways, such as their characterization of natural selection and fitness.
Deviation from mathematical expectation should be sharply distinguished from deviation from prediction. Brandon identifies drift with the former, but others, like Walsh et al. (2002) fail to sharply distinguish between prediction and expectation—as their quote at the end of the previous paragraph evinces, they cash out statistical error in terms of predictions in some places, but they also base drift on deviations from expectation in other places, e.g., “because the outcome diverges somewhat from expectation, there is drift too” (p. 464).
This is not a denial of determinism (though I take it that there is good scientific support for fundamental indeterminism). Instead, this is possible because ‘environment’ refers to a broader set of factors than those that happen to determine a particular outcome of a particular life. This is analogous to the observation that it is not inconsistent to hold that the (conditional) probability of a fair coin landing heads in a coin flipping machine is 0.5, whether or not the world is deterministic. See Sober (2010) for a discussion of this point.
Strictly speaking, the set of possible lives is infinite and the probability of each of the \(L_\mathrm{i}\) is zero. Thus it does not make sense to speak of differences in the probabilities of the various \(L_\mathrm{i}\). But because some of these lives will be effectively equivalent in terms of their outcomes, they can be bundled together in such a way that there is a finite number of bundles and each bundle has a non-zero probability. To not overly complicate the following discussion, I will simply speak of probabilities associated with particular possible lives, even though it is only the bundles that differ in their probabilities.
Describing this function precisely is a deep and difficult question, and one beyond the scope of the current discussion (see Pence and Ramsey forthcoming).
Under the label of ‘heterogeneity’ fall such things as differences in the timing, quality, and quantity of the offspring produced. The function on this heterogeneity will thus need to incorporate all of these facets of \(L_\mathrm{i}\) heterogeneity.
Note that I am here explicating organismic fitness and selection, not the fitness and selection of traits. These notions of fitness and selection are distinct (though related) and, for the sake of simplicity, I will not here discuss trait fitness/selection.
Similarly, flipping a set of coins with a ‘2’ on each side will tend to lead to a different tally from that of the same size set of coins with a ‘1’ on one side and a ‘3’ on the other, even though the expected tally is the same.
As with fitness, the task of specifying the nature of this heterogeneity and this function will be left to another paper.
By claiming that selection and driftability are in the same “ontological arena” I mearly mean that they are structurally the same, that they both consist in heterogeneity in the \(L_\mathrm{i}\).
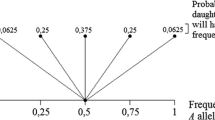
Because driftability includes germline mutations (such a mutation can be understood as a heritable transformation occuring along part of an \(L\)), the claim that \(\text{ drift}_\mathrm{o}\) cannot occur in an infinite population requires the assumption that the individuals in the population are not sui generis. Instead, each type in the infinite population must be represented by an infinite number of individuals. If this were not the case, there could be \(\text{ drift}_\mathrm{o}\) even in an infinite population. To see how this is possible, consider the probability of surviving extinction for a particular allele that arises in an individual. This probability is approximately \(2k\), where \(k\) is the advantage conferred by the allele (Haldane 1927). And since \(k\) is (or can be) constant over changes in population size, \(\text{ drift}_\mathrm{o}\) is not eliminated by an infinite population size (this requires the assumption that the offspring are drawn from a Poisson distribution—see Haldane 1927).
This implies that a rejection of the causal efficacy of dispositional properties would allow one to accept the definition of driftability and selection in terms of \(L_\mathrm{i}\) heterogeneity without holding that driftability or selection can cause evolution. Thus, a statisticalist could hold that the driftability framework is the right way to think about drift, but that while driftability quantifies and correctly locates the basis of drift in features of individual organisms, driftability is not a cause of \(\text{ drift}_\mathrm{o}\).
This way of quantifying fragility will have some odd implications. A glass that will almost certainly not break over its life but, if broken, will shatter into a billion pieces, will obtain a higher fragility value than a mug that always breaks, but always does so into a few pieces. If one is perturbed by these results, there are different ways to reduce the distribution to a single value. For example, using the mode of the distribution (instead of the mean) would reverse the rank ordering of the fitness values for the glasses and mugs, avoiding this counterintuitive result.
Arithmetic mean, it has been shown, is not a good metric for fitness in all situations. But more sophisticated metrics are available that are much more general (Pence and Ramsey, forthcoming).
Because selection and driftability could be independently intervened upon, the manipulability construal of causal explanation (Woodward 2005) would hold that selection and driftability can form the basis of independent, causal evolutionary explanations.
Direct access to the \(L_\mathrm{i}\) is of course impossible. But the heterogeneity in outcomes can be inferred either from (developmental, environmental, etc.) features affecting the organism’s life history, or from observing a sample of like organisms.
There is, however, one possible point of tension between the driftability framework and the PD, since the PD explicitly excludes drift from being an evolutionary “force.” I will not here lay out what I consider to be the necessary and sufficient conditions for something to qualify as a force, but the argument for driftability does take driftability to be a cause of evolutionary change. And, as suggested in Sect. 9 below, since driftability and selection are in the same ontological arena, it seems that if selection is a force, drift is too. If driftability were to be understood to be a force, then one could simply amend the PD by, for example, inserting ‘additional’ or ‘counterveiling’ before ‘evolutionary’ in condition (A).
References
Abrams, M. (2007). How do natural selection and random drift interact? In: C. Bicchieri & J. Alexander (Eds.), Proceedings of the 2006 biennial meeting of the Philosophy of Science Association. Part I: Contributed papers. Philosophy of Science, 74(5), 666–679.
Ariew, A., & Ernst, Z. (2009). What fitness can’t be. Erkenntnis, 71(3), 289–301.
Ariew, A., & Lewontin, R. C. (2004). The confusions of fitness. British Journal for the Philosophy of Science, 55, 347–363.
Beatty, J. H. (1984). Chance and natural selection. Philosophy of Science, 51, 183–211.
Beatty, J. H., & Finsen, S. (1989). Rethinking the propensity interpretation of fitness: A peek inside pandora’s box. In M. Ruse (Ed.), What the philosophy of biology is: Essays for David Hull (pp. 17–30). Dordrecht: Kluwer.
Bouchard, F., & Rosenberg, A. (2004). Fitness, probability and the principles of natural selection. British Journal for the Philosophy of Science, 55(4), 693–712.
Brandon, R. N. (1978). Adaptation and evolutionary theory. Studies in History and Philosophy of Science, 9(3), 181–206.
Brandon, R. N. (2005). The difference between selection and drift: A reply to Millstein. Biology and Philosophy, 20(1), 153–170.
Brandon, R. N. (2006). The principle of drift: Biology’s first law. Journal of Philosophy, 102(7), 319–335.
Brandon, R. N., & Carson, S. (1996). The indeterministic character of evolutionary theory: No ‘no hidden variables proof’ but no room for determinism either. Philosophy of Science, 63(3), 315–337.
Darwin, C. (1859). On the origin of species. London: John Murray.
Gildenhuys, P. (2009). An explication of the causal dimension of drift. British Journal for the Philosophy of Science, 60(3), 521–555.
Haldane, J. B. S. (1927). A mathematical theory of natural and artificial selection, part v: Selection and mutation. Proceedings of the Cambridge Philosophical Society (pp. 838–844). Oxford: Oxford University Press.
Haldane, J. B. S. (1954). The measurement of natural selection. Proceedings on IX International Congress Genetics, 1, 480–487.
Krimbas, C. B. (2004). On fitness. Biology and Philosophy, 19(2), 185–203.
McGraw, J. B., & Caswell, H. (1996). Estimation of individual fitness from life-history data. American Naturalist, 147(1), 47–64.
Matthen, M. (2009). Drift and “statistically abstractive explanation. Philosophy of Science, 76, 464–487.
Matthen, M., & Ariew, A. (2002). Two ways of thinking about fitness and natural selection. Journal of Philosophy, 99(2), 55–83.
McShea, D., & Brandon, R. N. (2010). Biology’s first law: The tendency for diversity and complexity to increase in evolutionary systems. Chicago: University of Chicago Press.
Mills, S. K., & Beatty, J. H. (1979). The propensity interpretation of fitness. Philosophy of Science, 46(2), 263–286.
Millstein, R. L. (2002). Are random drift and natural selection conceptually distinct? Biology and Philosophy, 17, 33–53.
Millstein, R. L. (2006). Natural selection as a population-level causal process. The British Journal for the Philosophy of Science, 57(4), 627–653.
Pence, C., Ramsey, G. (forthcoming). A new foundation for the propensity interpretation of fitness. British Journal for the Philosophy of Science.
Pfeifer, J. (2005). Why selection and drift might be distinct. Philosophy of Science, 72(5), 1135–1145.
Rosenberg, A. (1994). Instrumental biology or the disunity of science. Chicago: University of Chicago Press.
Sober, E. (1984). The nature of selection. London: MIT Press.
Sober, E. (2001). The two faces of fitness. In E. Sober (Ed.), Conceptual issues in evolutionary biology (pp. 25–38). London: MIT Press.
Sober, E. (2010). Evolutionary theory and the reality of macro-probabilities. In E. Ellery & J. H. Fetzered (Eds.), The place of probability in science (pp. 133–161). New York: Springer.
Stephens, C. (2004). Selection, drift, and the ’forces’ of evolution. Philosophy of Science, 71(4), 550–570.
Wagner, G. (2010). The measurement theory of fitness. Evolution, 64(5), 1358–1376.
Walsh, D. (2007). The pomp of superfluous causes: The interpretation of evolutionary theory. Philosophy of Science, 74(3), 281–303.
Walsh, D. M., Lewens, T., & Ariew, A. (2002). The trials of life: Natural selection and random drift. Philosophy of Science, 69(3), 452–473.
Woodward, J. F. (2005). Making things happen: A theory of causal explanation. Oxford: Oxford University Press.
Acknowledgments
This paper was the subject of a symposium at the 2011 meeting of the Pacific Division of the American Philosophical Association. I thank my commentators, Vadim Keyser, Mohan Matthen, and Sarah Roe for their insightful comments. This paper was also presented at the 2011 Workshop on Metaphysics of Microevolutionary Processes at the University of Alabama at Birmingham. I thank the organizer, Marshall Abrams, as well as the other attendees for their input. I also thank Robert Brandon, Yoichi Ishida, Bence Nanay, Charles Pence, and the anonymous reviewers for reading and commenting on earlier versions of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ramsey, G. Driftability. Synthese 190, 3909–3928 (2013). https://doi.org/10.1007/s11229-012-0232-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11229-012-0232-6