
Abstract
Drift is often characterized in statistical terms. Yet such a purely statistical characterization is ambiguous for it can accept multiple physical interpretations. Because of this ambiguity it is important to distinguish what sorts of processes can lead to this statistical phenomenon. After presenting a physical interpretation of drift originating from the most popular interpretation of fitness, namely the propensity interpretation, I propose a different one starting from an analysis of the concept of drift made by Godfrey-Smith. Further on, I show how my interpretation relates to previous attempts to make sense of the notion of expected value in deterministic setups. The upshot of my analysis is a physical conception of drift that is compatible with both a deterministic and indeterministic world.
Similar content being viewed by others


Notes
Note that this way of characterizing drift might not capture all the different meanings of drift used in the biological literature, but it captures some important meanings of it and is grounded in influential theoretical work (e.g., Grafen 2000; Rice 2004, 2008). For thorough reviews of the notion of drift see Plutynski (2007) and Millstein (2016).
Note that strictly speaking expected reproductive outputs are just one way to characterize propensities. In simple models propensities can be reasonably approximated with expected value (e.g., Mills and Beatty 1979). But doing so leads to a number of problems (see, for instance, Beatty and Finsen 1989). A number of authors have proposed new propensity interpretations of fitness (e.g., Brandon 1990; Ramsey 2006; Pence and Ramsey 2013) or some other interpretation (e.g., Abrams 2009) that attempt to solve these problems.
By “entity” I will mean any object of a population able to undergo evolution by natural selection (e.g., organism, gene, cell, group).
Note that some could consider that small differences in the environment affecting reproductive outputs are in some sense the “brute indeterministic facts about reality” even though the setup is deterministic. This will not be the interpretation given here since by definition indeterministic facts cannot occur in a deterministic setup. I recognize that epistemically one might want to take some facts as indeterministic even though the setup is deterministic, but the epistemic and the ontological questions should not be confused.
I am appealing here to the same kind of distinction made by Rosenberg (2001, pp. 537–538) between a statistical or indeterministic theory and an indeterministic world. Having a theory that is indeterministic does not imply that the world is and vice versa. Here the relevant distinction is that although a system might be deterministic, its input might not, and vice versa.
Although, as Godfrey-Smith recognizes, the terms “intrinsic” and “extrinsic” are controversial in philosophy, I will follow him in his view that the main idea behind the distinction can be useful. For more on this distinction see Weatherson (2014).
This consequence relies on the assumption that no asymmetric extrinsic properties (e.g., “being less tall than”) are involved in differences in reproductive outputs, in which case it would be impossible to have the same extrinsic property for all the members of a population. This assumption is of course far from any real biological case, but I use it merely to make a conceptual distinction.
I use here Brandon’s (1990) notion of selective environment as all the factors surrounding an organism (or more generally an entity) that differentially affect its reproductive output when compared to another organism.
This way of representing the environment in the context of evolutionary change is compatible with Abrams’s (2014) framework on environmental variation.
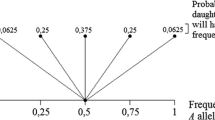
Suppose, for instance, a case with ten possible microstates present in equal proportions. It would imply that entities encounter each microstate with equal probability (0.1).
This explanation is to put in perspective the account proposed by Ramsey (2013), for whom drift results from the heterogeneity in the possible causes one entity can experience, leading to different reproductive outputs.
Thanks to Kim Sterelny for bringing this case to my attention.
According to Godfrey-Smith (2009, p. 63), this list of five parameters is incomplete and could include other features.
For our purposes in this article this is equivalent to realized reproductive output.
Specifically, the Breeder’s equation tells us that the response to selection R is equal to a selection differential S multiplied by heritability h 2 so that R = S × h 2.
I write “usually” because some plants need fire for their seeds to germinate, for instance.
This example is mistakenly attributed to Scriven (1959), who was rather talking of individuals sitting where a bomb or a tree falls.
Note here that this problem is not encountered in Godfrey-Smith’s initial account. This is because in his account of drift, the parameter C introduces an element of randomness.
Note that the contrapositive is not true: something unpredictable might not necessarily be random.
Note however that some might want to define drift more broadly than it is classically done and include cases of correlated responses as cases of drift.
Note that by “unpredictable” I am referring here to a single outcome. In the case of drift this would amount to the evolutionary success of a single allele. At the population level it is quite predictable that a population will exhibit drift.
References
Abrams M (2009) The unity of fitness. Philos Sci 76(5):750–761
Abrams M (2012a) Mechanistic probability. Synthese 187(2):343–375
Abrams M (2012b) Mechanistic social probability: how individual choices and varying circumstances produce stable social patterns. In: Kincaid H (ed) The Oxford handbook of philosophy of social science. Oxford University Press, Oxford, pp 184–226
Abrams M (2014) Environmental grain, organism fitness, and type fitness. In: Barker G, Desjardins E, Pearce T (eds) Entangled life. Springer, Netherlands, pp 127–151
Beatty JH, Finsen SK (1989) Rethinking the propensity interpretation: a peek inside Pandora’s box. In: Ruse M (ed) What the philosophy of biology is: essays dedicated to David Hull. Kluwer Publishers, Dordrecht, pp 17–30
Bouchard F, Rosenberg A (2004) Fitness, probability and the principles of natural selection. Br J Philos Sci 55(4):693–712
Bourrat P (2015a) Distinguishing natural selection from other evolutionary processes in the evolution of altruism. Biol Theory 10(4):1–11. doi:10.1007/s13752-015-0210-6
Bourrat P (2015b) How to read “heritability” in the recipe approach to natural selection. Br J Philos Sci 66(4):883–903
Bourrat P, Lu Q (2016) Dissolving the missing heritability problem. Philos Sci
Brandon RN (1978) Adaptation and evolutionary theory. Stud Hist Philos Sci Part A 9(3):181–206
Brandon RN (1990) Adaptation and environment. Princeton University Press, Princeton
Brandon RN, Carson S (1996) The indeterministic character of evolutionary theory: no ‘no hidden variables proof’ but no room for determinism either. Philos Sci 63(3):315–337
Downes SM (2009) Moving past the levels of selection debates. Biol Philos 24(5):703–709
Eagle A (2004) Twenty-one arguments against propensity analyses of probability. Erkenntnis 60(3):371–416
Falconer DS (1981) Introduction to quantitative genetics. Longman, New York
Gillespie JH (2004) Population genetics: a concise guide. Johns Hopkins University Press, Baltimore
Glymour B (2001) Selection, indeterminism, and evolutionary theory. Philos Sci 68(4):518–535
Godfrey-Smith P (2007) Conditions for evolution by natural selection. J Philos 104(10):489–516
Godfrey-Smith P (2009) Darwinian populations and natural selection. Oxford University Press, New York
Grafen A (2000) Developments of the Price equation and natural selection under uncertainty. Proc R Soc Lond B Biol Sci 267(1449):1223–1227
Hájek A (2012) Interpretations of probability. Stanford Encyclopedia of Philosophy. http://plato.stanford.edu/archives/win2012/entries/probability-interpret/
Hamilton M (2009) Population genetics. Wiley-Blackwell, Oxford
Lu Q, Bourrat P (2016) The evolutionary gene and the extended evolutionary synthesis. Br J Philos Sci
Lynch KE, Bourrat P (2016) Interpreting heritability causally. Philos Sci. doi:10.1086/688933
Mills SK, Beatty JH (1979) The propensity interpretation of fitness. Philos Sci 46(2):263–286
Millstein RL (2002) Are random drift and natural selection conceptually distinct? Biol Philos 17(1):33–53. doi:10.1023/a:1012990800358
Millstein RL (2003) Interpretations of probability in evolutionary theory. Philos Sci 70(5):1317–1328. doi:10.1086/377410
Millstein RL (2016) Genetic rift. In: Zalta EN (ed) The Stanford encyclopedia of philosophy, Fall 2016. http://plato.stanford.edu/archives/fall2016/entries/genetic-drift/
Okasha S (2006) Evolution and the levels of selection. Oxford University Press, New York
Pence CH, Ramsey G (2013) A new foundation for the propensity interpretation of fitness. Br J Philos Sci 64(4):851–881
Pfeifer J (2005) Why selection and drift might be distinct. Philos Sci 72(5):1135–1145
Plutynski A (2007) Drift: a historical and conceptual overview. Biol Theory 2(2):156–167. doi:10.1162/biot.2007.2.2.156
Ramsey G (2006) Block fitness. Stud Hist Philos Sci Part C 37(3):484–498
Ramsey G (2013) Driftability. Synthese 190(17):3909–3928. doi:10.1007/s11229-012-0232-6
Rice SH (2004) Evolutionary theory: mathematical and conceptual foundations. Sinauer, Sunderland
Rice SH (2008) A stochastic version of the Price equation reveals the interplay of deterministic and stochastic processes in evolution. BMC Evol Biol 8:262. doi:10.1186/1471-2148-8-262
Rosenberg A (2001) Discussion note: indeterminism, probability, and randomness in evolutionary theory. Philos Sci 68(4):536–544
Rosenthal J (2010) The natural-range conception of probability. In: Ernst G, Hüttemann A (eds) Time, chance, and reduction: philosophical aspects of statistical mechanics. Cambridge University Press, Cambridge, pp 71–90
Scriven M (1959) Explanation and prediction in evolutionary theory. Science 130:477–482
Sober E (1984) The nature of selection. MIT Press, Cambridge
Sober E (2001) The two faces of fitness. In: Singh RS, Krimbas CB, Paul DB, Beatty J (eds) Thinking about evolution: historical, philosophical, and political perspectives. Cambridge University Press, Cambridge
Sober E (2010) Evolutionary theory and the reality of macro-probabilities. In: Eels E, Fetzer JH (eds) The place of probability in science. Springer, Dordrecht, pp 133–161
Stamos DN (2001) Quantum indeterminism and evolutionary biology. Philos Sci 68(2):164–184
Stephens C (2004) Selection, drift, and the ‘forces’ of evolution. Philos Sci 71(4):550–570. doi:10.1086/423751
Strevens M (2011) Probability out of determinism. In: Beisbart Hartmann (ed) Probabilities in physics. Oxford Univeristy Press, New York, pp 339–364
Strevens M (2016) The reference class problem in evolutionary biology: distinguishing selection from drift. In: Ramsey G, Pence CH (eds) Chance in Evolution. University of Chicago Press, Chicago, pp 145–175
Weatherson B (2014) Intrinsic vs. extrinsic properties. Stanford Encyclopedia of Philosophy. http://plato.stanford.edu/archives/fall2014/entries/intrinsic-extrinsic/
Weber M (2001) Determinism, realism, and probability in evolutionary theory. Philos Sci 68(3):S213–S224
Acknowledgments
I am thankful to Mark Colyvan, Paul Griffiths, Aidan Lyon, Maureen O’Malley, Charles Pence, Arnaud Pocheville, and Kim Sterelny for their comments on a previous version of the manuscript. This research was supported under the Australian Research Council’s Discovery Projects funding scheme (Project Number DP150102875).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bourrat, P. Explaining Drift from a Deterministic Setting. Biol Theory 12, 27–38 (2017). https://doi.org/10.1007/s13752-016-0254-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13752-016-0254-2