
Abstract
Biosemiotics argues that “sign” and “meaning” are two essential concepts for the explanation of life. Peircean biosemiotics, founded by Tomas Sebeok from Peirce’s semiotics and Jacob von Uexkül’s studies on animal communication, today makes up the mainstream of this discipline. Marcello Barbieri has developed an alternative account of meaning in biology based on the concept of code. Barbieri rejects Peircean biosemiotics on the grounds that this discipline opens the door to nonscientific approaches to biology through its use of the concept of “interpretation.” In this article, it is noted that Barbieri does not adequately distinguish among Peirce’s semiotics, Peircean biosemiotics, and “interpretation-based” biosemiotics. Two key arguments of Barbieri are criticized: his limited view of science and his rejection of “interpretation-based” biosemiotics. My argument is based on tools taken from a different approach: Robert Rosen’s relational biology. Instead of “signs” and “meanings,” the study begins in this case from the “components” and “functions” of the organism. Rosen pursues a new definition of a law of nature, introduces the anticipatory nature of organisms, and defines the living being as a system closed to efficient cause. It is shown that Code Biosemiotics and Peircean biosemiotics can share a common field of study. Additionally, some proposals are suggested to carry out a reading of Rosen’s biology as a biosemiotic theory.

Modified from Rosen (1991, p. 60, Fig. 3H.2)

Modified from Rosen (2012, p. 75, Fig. 2.2)


Modified from Rosen (2012, p. 76, Fig. 2.3)

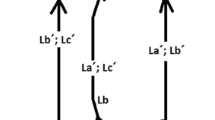
Modified from Rosen (1991, p. 53, Fig. F.2)


Modified from Rosen (1991, p. 251, Fig. 10C.6)
Similar content being viewed by others

Change history
22 August 2018
In the ‘Barbieri’s Concept of Mechanisms’ section on page 12 of above mentioned article.
Notes
The term “semiosis” describes the process of producing and responding to signs. Semiotics is the study of semiosis.
Peirce writes: “I define a sign as anything which is so determined by something else, called its Object, and so determines an effect upon a person, which effect I call its interpretant, that the later is thereby mediately determined by the former” (1998, p. 478).
Robert Rosen was also a physicist and mathematician, and in these disciplines the beauty of the theories is a criterion that counts.
Atkin (2013) provides an interesting overview of Peirce’s theory of signs.
Peirce used different terms for the signifying element including “sign,” “representamen,” “representation,” and “ground.” John Deely (1990) introduced the notion of “sign vehicle” to interpret and develop Peirce’s semiotics.
See Brier (2008) for a detailed study of Peircean biosemiotics.
Hoffmeyer defines semiotic freedom as “the ‘depth of meaning’ that an individual or a species is capable of communicating” (1996, p. 62).
Over four decades, Barbieri has been developing a theoretical framework for the study of biology. What this author began by calling Semantic Biology or Biosemantics (Barbieri 1985) was then changed to Code Biosemiotics and finally to Code Biology. These name changes correspond to three stages, which do not respond to changes in his proposals, but in his relations with other lines of research: a first stage of work independent of other schools that also consider semiosis to be the key to the study of biology; a second stage in which an attempt was undertaken to unify all schools under the common framework of Biosemiotics; and a third in which Barbieri and his collaborators took an independent path again, rejecting the prevailing line in Biosemiotics, considering it based on interpretation and often far from scientific work. In a letter to the editor of Biological Theory, published in 2014, Barbieri explained the reasons that led him to join the Biosemiotic movement in 2001 and to abandon it in 2012 to establish Code Biology as an independent research field.
The application of (conventionally established) rules determines which pairs of molecules are related. This relationship gives each molecule of the first set a meaning in the second set, and the expression of that meaning defines biological specificity.
In the case of the genetic code, the correspondence between codons and amino acids can only be the result of conventional rules. The genetic code had to appear at the same time as the translation apparatus. The result is a codemaker, a machine to produce proteins from a template and a code.
Barbieri classifies life into three worlds. The first world is based only on organic semiosis. Its mechanism is coding, and its codes are organic codes. The second world incorporates animal semiosis, which is characterized by two mechanisms (coding and interpretation) and two types of codes (organic and neural). The third world adds human semiosis. It is based on three mechanisms (coding, interpretation, and language) and three types of codes (organic, neural, and language).
According to Rosen (1991), a natural system N can have analytical and synthetic models. Analytical models are tied to the idea of the Cartesian (or direct) product of quotient spaces, while synthetic models are linked to the idea of the direct sum of subspaces.
Consider a property of system N, embodied in a model M, which is smaller than the maximum model, and the maximum model can be decomposed as the direct sum of two summands, with M being one of them. If this process can be performed for all of the properties of N, then we say that N can be fractionated, and its properties are fractionables.
“I call a system which is not simple ‘complex’. Complex systems cannot be exhausted by any finite number of simple (mechanical) models; they cannot be described as software to a ‘machine’” (Rosen 2006, p. 21).
A definition is said to be impredicative if it invokes the set being defined. It is a property that important mathematicians and logicians, especially Russell, wanted to leave out of their disciplines, but it has long been proved that elimination leads to having to abandon much of mathematics.
Unlike Barbieri, Rosen argues that only one mechanism of evolution is necessary: natural selection.
Taking Aristotelian causes as a reference, we can consider that A is the material cause of B (each element of A is the material cause of an element of B), whereas f is the efficient cause of the occurrence of the flow, i.e., the efficient cause of B (of each element of B).
The first edition of Anticipatory Systems dates from 1985.
“β/B” has been used instead, where “B” appears in Rosen’s original diagram, to avoid the usual error of identifying β with B, or with an element b of set B. According to Rosen (1991), β is the inverse of an operator constructed from b ∈B.
See Rosen’s quotation in the next section.
Nicholson shows the problems generated by not separating the three meanings, even among the new mechanists, quoting an example (Craver and Darder 2005, p. 234) in which “(T)he concept is used in different senses, sometimes even in the same passage (Nicholson 2012, p. 154).”
As Nicholson (2012, p. 154) explains, “The new mechanism discourse is not committed to a mechanistic worldview, nor does it prescribe a mechanistic approach in biology.”
The Ising model is usually considered a mathematical model of ferromagnetism, although Rosen points out: “[T]he Ising model is a metaphor for phase transitions, and not a ‘model’ in our sense” (Rosen 2012, p. 179; italics in original).
It has been argued that Rosen’s assertions on non-simulability of (M, R)-systems have not been refuted. In any case, although the non-simulability of the (M, R)-systems was refuted, as long as the non-simulability of the organisms is maintained, the claim that an organism is not a mechanism, and, therefore, the science of biology must be broader than the science of mechanisms remains valid. This would be sufficient to support our criticism of Barbieri’s limited vision of science, and to defend the compatibility of Barbieri’s biosemiotics with Peircean biosemiotics.
Note that this last sentence, which here applies to Rosen, is literally included in Brier’s explanation of Hoffmeyer (Brier 2008, p. 49).
Pattee’s biosemiotics also requires memory. Biological constraints can only occur in individuals with memory maintained by natural selection. Only individuals with memory-based control can learn from the environment and evolve (Pattee and Kull 2009).
“(H)is emphasis on ‘relational biology’ depended on semiotic rather than material relations. Rosen and I were friends for many years beginning with our studies of hierarchy theory in the 1960s. To us, hierarchies, like measurement, are also dependent on semiotic distinctions because hierarchical levels are recognized by the necessity of different descriptions” (Pattee and Kull 2009, p. 327).
References
Alberts B, Johnson A, Lewis J et al (2007) Molecular biology of the cell, 5th edn. Garland, New York
Atkin A (2013) Peirce’s theory of signs. In: Zalta EN (ed) The Stanford encyclopedia of philosophy (summer 2013 edition). https://plato.stanford.edu/archives/sum2013/entries/peirce-semiotics/
Barbieri M (1981) The ribotype theory on the origin of life. J Theor Biol 91:545–601
Barbieri M (1985) The semantic theory of evolution. Harwood Academic Publishers, London
Barbieri M (2003) The organic codes: an introduction to semantic biology. Cambridge University Press, Cambridge
Barbieri M (2008) Biosemiotics: a new understanding of life. Naturwissenschaften 95:577–599
Barbieri M (2009) A short history of biosemiotics. Biosemiotics 2:221–245
Barbieri M (2014) From biosemiotics to Code Biology. Biol Theory 9(2):239–249
Barbieri M (2015) Code Biology: a new science of life. Springer, Cham
Bechtel W (2006) Discovering cell mechanisms: the creation of modern cell biology. Cambridge University Press, Cambridge
Bechtel W (2007) Biological mechanisms: organized to maintain autonomy. In: Boogerd F, Bruggerman F, Hofmeyr JH, Westerhoff HV (eds) Systems biology: philosophical foundations. Elsevier, Amsterdam, pp 269–302
Berridge M (1985) The molecular basis of communication within the cell. Sci Am 253:142–152
Brier S (2008) The paradigm of Peircean biosemiotics. Signs 2:20–81
Cárdenas ML, Letelier JC, Gutierrez C et al (2010) Closure to efficient causation, computability and artificial life. J Theor Biol 263(1):79–92
Cornish-Bowden A, Piedrafita G, Morán F et al (2013) Simulating a model of metabolic closure. Biol Theory 8(4):383–390
Deely J (1990) Basis of semiotics. Indiana University Press, Bloomington
Emmeche C, Kull K, Stjernfelt F (2002) Reading Hoffmeyer, rethinking biology. Tartu Semiotics Library Vol. 3. Tartu University Press, Tartu
Favareau D (2006) The evolutionary history of biosemiotics. In: Barbieri M (ed) Introduction to biosemiotics: the new biological synthesis. Springer, Berlin, pp 1–67
Frege G (1892) Über Sinn und Bedeutung. Zeitschrift für Philosophie und philosophische Kritik 100:25–50. English translation in Geach P, Black M (eds) (1952) Translations from the philosophical writings of Gottlob Frege. Blackwell, Oxford
Hoffmeyer J (1996) Signs of meaning in the universe. Indiana University Press, Bloomington
Hoffmeyer J (1997) The swarming body. In: Rauch I Carr GF (eds) Semiotics around the world: synthesis in diversity. Proceedings of the 5th congress of the International Association for Semiotic Studies. Berkeley 1994. Mouton de Gruyter, Berlin, pp 937–940
Hoffmeyer J (2009) Epilogue: biology is immature biosemiotics. In: Deely J, Shrocci L (eds.) Proceedings of the 33rd annual meeting of the Semiotic Society of America. Legas Publishing, Ottawa, pp 927–942
Kauffman S (2000) Investigations. Oxford University Press, New York
Kull K (2001) Jakob von Uexküll: a paradigm for biology and semiotics. Semiotica 134(1/4). Mouton de Gruyter, Berlin
Moreno A, Mossio M (2015) Biological autonomy: a philosophical and theoretical enquiry. Springer, Dordrecht
Mossio M (2013) Closure, causal. In: Dubitzky W, Wolkenhauer O, Cho K-H, Yokota H (eds) Encyclopedia of systems biology. Springer, New York, pp 415–418
Mossio M, Longo G, Stewart J (2009) A computable expression of closure to efficient causation. J Theor Biol 257(3):489–498
Nicholson D (2012) The concept of mechanism in biology. Stud Hist Philos Biol Biomed Sci 43(1):152–163
Pattee HH (1968) The physical basis of coding and reliability in biological evolution. In: Waddington CH (ed) Towards a theoretical biology, vol 1. Edinburgh University Press, Edinburgh, pp 67–93
Pattee HH (1972) Laws and constraints, symbols and languages. In: Waddington CH (ed) Towards a theoretical biology, vol 4. Edinburgh University Press, Edinburgh, pp 248–258
Pattee HH (2007) Laws, constraints and the modeling relation—history and interpretations. Chem Biodivers 4:2272–2295 2007)
Pattee HH, Kull K (2009) A biosemiotic conversation: between physics and semiotics. Sign Syst Stud 37(1/2):311–331
Peirce CS (1931–1936) The collected papers, vol. 1–6. In: Hartshorne C, Weiss P (eds) Harvard University Press, Cambridge
Peirce CS (1998) The essential Peirce, vol. 2. Peirce Edition Project. Indiana University Press, Bloomington
Posner R, Robering K, Sebeok TA (1997) Semiotik/semiotics: a handbook on the sign-theoretical foundations of nature and culture, vol 1. Walter de Gruyter, Berlin
Rashevsky N (1954) Topology and life: in search of general mathematical principles in biology and sociology. Bull Math Biophys 16:317–348
Rosen R (1991) Life itself. A comprehensive enquiry into the nature, origin and fabrication of life. Columbia University Press, New York
Rosen R (2006) Autobiographical reminiscences of Robert Rosen. Axiomathes 16(1–2):1–23. https://doi.org/10.1007/s10516-006-0001-6
Rosen R (2012) Anticipatory systems. Philosophical, mathematical, and methodological foundations, 2nd edn. Springer, New York. [1st edn, 1985, Pergamon Press, Ontario]
Sebeok TA (1963) Communication among social bees; porpoises and sonar; man and dolphin. Language 39:448–466
Sebeok TA (1972) Perspectives in zoosemiotics. Mouton, The Hague
Sebeok TA (2001) Biosemiotics: its roots, proliferation, and prospects. In: Kull K (ed) Jakob von Uexküll: a paradigm for biology and semiotics. Semiotica 134(1/4):61–78
Uexküll J von (1928) Theoretische biologie, 2nd edn. Julius Springer, Berlin
Varela FJ (1979) Principles of biological autonomy. Elsevier, New York
Acknowledgments
I thank Cristian Saborido and two anonymous reviewers for their valuable comments on previous versions of this article. Deborah Klosky deserves special thanks for her editorial assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
The original version of this article was corrected: In the ‘Barbieri’s Concept of Mechanisms’ section on page 12 of above mentioned article the text erroneously reads Rosen, quoting Alberts et al. (2007), highlights…’. The correct text should read ‘Barbieri, quoting Alberts et al. (2007), highlights…’.
Rights and permissions
About this article

Cite this article
Vega, F. A Critique of Barbieri’s Code Biology Through Rosen’s Relational Biology: Reconciling Barbieri’s Biosemiotics with Peircean Biosemiotics. Biol Theory 13, 261–279 (2018). https://doi.org/10.1007/s13752-018-0302-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13752-018-0302-1